 |
|
МЕНЮ
|
Реферат: Зависимость интенсивности дыхания растительных продуктов от температурыРЕЖИМ ХРАНЕНИЯ ОВОЩЕЙ И КАРТОФЕЛЯ
Таблица 1(а). Осенью, когда температура воздуха бывает повышенной, на ночь для создания сквозняка открывают все окна, вытяжные трубы, люки, двери, а днем их закрывают, чтобы сохранить запас холода. Иногда в погребах отмечается высокая влажность воздуха. Ее признаками служат затхлый спертый воздух, ощущение сырости, появление плесени и конденсированной влаги на потолке, стенах, стеллажах, ящиках. Чтобы понизить чрезмерную влажность воздуха, одновременно открывают двери и отдушины. Можно поставить в подвал ящик с негашеной известью, солью или древесным углем — эти материалы впитывают воду. При похолоданиях утепляют крышу. С декабря наступает опасность промерзания хранилища. К этому времени двери, а при необходимости и стены обивают матами и рогожей. Вентиляционные трубы, кроме нужных для работы, плотно закрывают соломой или другими материалами. 8)БЫСТРОЕ ЗАМОРАЖИВАНИЕ Цель быстрого замораживания, как и любого другого консервирующего процесса, свести на нет или, по крайней мере, замедлить реакции, ухудшающие качество продуктов и в конечном итоге делающие их непригодными к употреблению. Практически можно говорить только о том, что же преобладает в применяемом процессе. По этому решающему действию или характеру процесса его относят к той или иной группе. Любой процесс консервирования тем лучше, чем меньшие изменения он вызывает в продуктах в сравнении с их первоначальными свойствами и чем более длительный срок хранения он обеспечивает. В настоящее время из применяемых в промышленных масштабах методов консервирования продуктов процессы холодильной обработки и замораживания лучше всего удовлетворяют упомянутым выше требованиям, поскольку, с одной стороны, они по отношению к другим процессам консервирования вызывают более слабые изменения свойств продуктов, а с другой—обеспечивают достаточно долгую для практических целей сохраняемость пищи. Процесс консервирования путем холодильной обработки занимает особое место еще и потому, что после охлаждения или замораживания продукты можно сохранять с помощью любого другого процесса консервирования и, наоборот, продукты, законсервированные с помощью других методов, хранятся дольше всего только в охлажденном состоянии; Процессы, ухудшающие качество продуктов, или процессы, приводящие в конечном итоге к их порче, развиваются в комплексных условиях. Различают физические, биохимические и биологические процессы порчи продуктов. Холодильная обработка независимо от характера обрабатываемого продукта действует на эти процессы с определенной закономерностью. Из физических процессов порчи к самым основным относятся процессы усушки продуктов или потери массы. Они вызывают не только количественные изменения (уменьшение массы), но и могут привести к ухудшению качества. Потеря массы продуктов подчинена следующей закономерности: ΔS = Βa(Pп-φp) где Δ S — теряемое продуктами количество воды за единицу времени, г/ч; Β — коэффициент испарения воды, г/(Н-ч); А—площадь поверхности продуктов, м2; рп — давление водяных паров при температуре поверхности продуктов, Па; φ — относительная влажность воздуха в виде десятичной дроби; р—давление насыщенных водяных паров при температуре окружающей среды, Па. При данной поверхности продуктов и данной величине коэффициента испарения потеря массы зависит от величин Pп, φр и р. Если одни и те же продукты с равновесным паросодержанием, например 95%, хранить при различных температурах в помещении с относительной влажностью 80%, то величина Рп—φр будет изменяться следующим образом (при условии, что температура продуктов равна температуре окружающего пространства):
Таким образом, при прочих равных условиях потеря массы при температуре 40° С примерно в 100 раз больше, чем потеря массы при —20° С, т. е. низкие температуры значительно уменьшают степень усушки продуктов. Мы говорили уже о том, что потерю массы можно уменьшить с помощью соответствующей упаковки, но даже в этом случае остается явление так называемой внутриупаковочной потери массы. Биохимические процессы порчи играют исключительно важную роль. Сюда относятся вызванные ферментами катализные процессы распада, включая даже микробиологические процессы порчи, поскольку ясно, что и эти процессы при ближайшем рассмотрении тоже носят биохимический характер. При высоких температурах химические изменения вещества проходят очень быстро, а скорость реакций высока, при низких температурах эти изменения часто едва заметны. Понижение температуры приводит к замедлению постоянных химических реакций, т. е. к уменьшению их скорости. Этот факт выражается законом Аррениуса, который связывает абсолютную температуру Т и логарифм коэффициента химической реакции следующим соотношением: logK=a—b/T. Коэффициенты а и b уравнения Аррениуса не зависят от температуры и находятся экспериментальным путем для каждой кон- j кретной реакции. Приведенное соотношение дает возможность вы- i числить скорость химической реакции при любой температуре. На основании этих расчетов и экспериментальных данных можно сделать вывод, что скорость наиболее простых (мономолекулярных, ; бимолекулярных) реакций при повышении температуры на 10° С увеличивается в 2—3 раза и соответственно понижение температуры на 10° С уменьшает ее до половины или до одной трети. На основании такой закономерности легко видеть, что скорость химических и биохимических реакций сильно замедляется с понижением температуры. Предположим, начальная скорость какой-нибудь реакции v при понижении температуры на 10° С уменьшилась до '/з своей величины, т. е. стала равна v/3. Если температура уменьшится теперь еще на 10° С, то скорость станет равной уже v/(3-3)=v/32 начальной скорости. Рассуждая подобным образом, увидим, что скорость уменьшается по степенному закону: v/33, v/3…v/з. Показатель степени п равен десятой части разности температур реакции. Например, если скорость реакции при 40ْС равна v, то при температуре —20° С она составит уже v/729. Интересно, однако, заметить, что биохимические реакции проходят на основании этого закона даже при очень низких температурах, только их скорость становится исключительно малой. Степень снижения скорости биохимических реакций при уменьшении температуры на 10°С, следуя ван Гоффу, принято обозначать Qio. Степенью Qio принято, однако, описывать снижение скорости не только какой-нибудь одной определенной реакции. Эту величину используют также для характеристики снижения скорости всей совокупности биохимических реакций в целом. Например, поскольку сохраняемость зависит главным образом от действия совокупности биохимических реакций, то срок хранения также может характеризоваться величиной Qio. Куприянов установил для различных видов пищи увеличение времени ее сохраняемости при изменении температуры на 10° С. Qio = где Qio—температурный коэффициент сохраняемости; длительность хранения при t=10°C; —продолжительность хранения при t С. Как установил Куприянов, при низкотемпературной хранении быстрозамороженных овощей величина Qio равна 2, нежнрной рыбы—3,1, а птицы—4. Интересная картина получается, если срок хранения какого-нибудь продукта, определенный различными авторами (например, продолжительность хранения говядины или свинины), отложить на графике зависимости от температуры в температурной области ниже и выше точки замерзания (рис. ).
Проделав соответствующие расчеты для значения Qio в области температур выше и ниже точки замерзания, мы получим следующие величины:
Говядина выше точки замерзания » ниже » » Свинина выше точки замерзания » ниже » » Видно, что при температурах ниже точки замерзания продолжительность хранения увеличивается быстрее при уменьшении температуры, чем в температурной области выше точки замерзания. Таким образом, после перехода через точку замерзания не только уменьшается скорость биохимических реакций, но вступает в силу и новый фактор, влияющий на удлинение срока сохраняемости продуктов. Таким фактором является вымерзание находящейся в продуктах воды (табл. ).
Снижение температуры оказывает многостороннее и комплексное воздействие на биологические процессы. Ниже мы будем рассматривать этот вопрос более подробно. Сейчас только заметим, что применение низких температур приводит к приостановлению и замедлению биологических процессов. С другой стороны, при низких температурах эти процессы могут прекратиться вообще, что вызывает иногда вредные, а иногда полезные изменения. Во время быстрого замораживания таких влагосодержащих веществ, как пищевые продукты, значительная часть воды становится льдом. Поэтому применение низких температур в подобных процессах консервирования обязательно сопровождается явлением замерзания. Следует, однако, упомянуть, что все-таки есть и такие процессы, где применение низких температур не сопровождается вымерзанием влаги из обрабатываемого продукта. Явление вымерзания удается исключить, когда точка замораживания продуктов сильно понижается с помощью введения в них каких-нибудь веществ, например, сахара или соли. Тогда температура хранения может быть ниже температуры замерзания. Другой пример тому—наличие связанной воды в материале, подвергаемом процессу быстрого замораживания. Связанная вода не вымерзает даже при очень низких температурах, например даже при вымерзании бактериальных спор. Так как процесс замерзания обычно играет важную роль при быстром замораживании продуктов в изменении их качества, мы познакомимся с ним более подробно. 9) ЗАМОРАЖИВАНИЕ КЛЕТОК Как животные, так и растительные клетки можно заморозить и в живом, и в мертвом состоянии. Замораживаются отдельные клетки в суспензии и клетки, образующие структуру тканей. Однако для всех этих случаев можно установить и одинаково справедливые общие закономерности. Как известно, химический состав клеток сложен. С точки зрения замораживания главную роль играет поведение клеточных белковых соединений. Благодаря своим физическим свойствам белки относятся к коллоидам. Растворяясь в воде, они образуют гидрогели и гидрозоли. Та часть воды, активность которой в гидрозолях и гидрогелях ниже нормальной, называется связанной водой, а остальное водосодержание—свободной водой. Под действием таких внешних воздействий, как изменение температуры, золи переходят в гели и наоборот. Изменения в клетках при их охлаждении начинают происходить уже вблизи точки замерзания. Вязкость клеточной протоплазмы возрастает, образуется гель, а содержащиеся в клетках липиды вымерзают. Существенные изменения, однако, начинаются после замерзания некоторого количества воды клеточной влаги. Замерзание клеточного коллоида обычно начинается так же, как и замерзание истинного раствора. Здесь тоже возникает явление понижения температуры замерзания, а из коллоидного раствора вымерзает только чистая вода. Вследствие вымерзания воды остаточная концентрация раствора возрастает, и температура его замерзания подобно истинному раствору уменьшается. Чем ниже температура клеток, тем больше из них вымерзает воды, но благодаря коллоидному характеру системы связанная вода обычно не замерзает. Так, по данным Морана, в 2%-ном растворе желатина примерно 35% воды не вымораживается даже при температуре жидкого воздуха (—196° С). На рис. температура t соответствует точке замерзания. Ее значение зависит от концентрации растворенных в клеточной влаге веществ. При понижении температуры и дальнейшем отборе тепла некоторая часть воды клеточной влаги вымерзает, но жизнеспособность организма пока не изменяется и не исчезает. Это явление справедливо даже для самых общих случаев. Такое состояние продолжается до температуры t. Говоря о вымерзании воды на участке t1—t2, мы имели в виду биохимически (и даже биологически) свободную воду. На этом участке скорость замерзания не играет особой роли. Случается даже так, что при медленном замерзании биологическая обратимость оказывается лучшей и большее количество клеток остается в живом состоянии. После оттаивания клеточный обмен веществ снова возобновляется. Тут, однако, становится важным фактор времени, в течение которого клетки находились при температуре t1 до tг, и то, каким образом происходило оттаивание. Значение температуры t2, или границы биологической обратимости, неоди- наково для разных клеток и клеточных структур. Биологическая обратимость при замораживании или полное восстановление жизненных функций после замораживания характерно только для некоторых пород рыб (карасей), некоторых растений и обычно структур более низкого порядка и организации. При охлаждении ниже температуры t2 процесс вымерзания воды продолжается и, хотя клетки и клеточные структуры отмира-
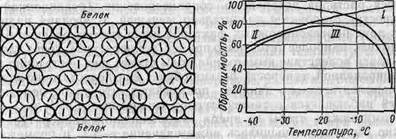
ют, вола вымерзает из коллоидного раствора обратимым образом Вымерзшее количество воды до температуры t3 полностью представлено коллоидно свободной водой. У фруктов температуры t2 и t3 находятся обычно очень близко друг от друга, т. е. прекращений жизнедеятельности практически сопровождается наступлением коллоидной необратимости процесса. Ниже температуры tз начинает замерзать связанная коллоидная вода. Коллоидные частички которые до этого были надежно отделены друг от друга оболочкой гидратов, теперь могут сближаться настолько, что различны силы притяжения склеивают мицеллы. Высвобожденные связи, на которых раньше находились молекулы воды, теперь действуют друг на друга так, что при взаимодействии некоторых главных соединений коллоидных частичек могут произойти химические изменения и коллоид может денатурироваться. Если позже к местам этих связей подойдут молекулы воды, то они не смогут взаимодействовать с ними. Исследования Риделя показали, что даже если температура клеток понизится ниже температуры t3, то и тогда вся вода не вымерзнет из растворов, т. е. содержащуюся в клетках воду невозможно выморозить полностью. Линг (1968) объясняет состояние связанной воды наличием на| молекулярной поверхности вещества, образующего коллоидный раствор, многослойного поляризованного покрытия из водяных молекул (рис. ). Свойства поляризованных водных молекул (их точка замерзания, способность растворять различные вещества и т.д.) отличаются от свойств нормальных неполяризованных молекул. Можно предположить, что полное влагосодержание живых клеток состоит именно из таких поляризованных молекул. Уменьшение обратимости коллоидов по мере вымерзания воды ведет к уплотнению остаточного раствора. Водные растворы, образующие клеточную влагу, в зависимости от вида продуктов со-
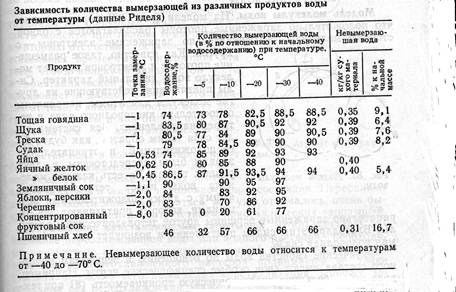
Рис. 91. Расположение слоев поляризованных молекул воды между молекулами белка. Черточки внутри кружков, обозначающих молекулы воды, показывают направления дипольных моментов. Рис. 92. Влияние температуры на обратимость замерзания. Кривая / показывает, что уменьшение температуры и увеличение скорости замерзания благоприятно действуют на обратимость, а кривая // показывает, что снижение температуры вследствие концентрации клеточной влаги действует на обратимость неблагоприятным образом. Характеризующая суммарное воздействие этих процессов кривая /// получается суперпозицией кривых / и //. держат соли, кислоты, сахара, коллоидные белки и т. д. Концентрация образующих электролиты или находящихся в диссоциированном состоянии солей и кислот вызывает коагуляцию или денатурирование белков. Во время замораживания недостаточно амортизированных систем может изменяться величина рН. Так, например, рН овощей: при замораживании может уменьшаться, а рН мяса увеличиваться. Эти данные указывают на то, что во время замораживания величина рН может достичь изоэлектрической точки выпадения белков или приблизиться к ней. Изоэлектрическая точка глютенина и яичного белка при рН 4,8, оксигемоглобина—при рН 6,8. С другой стороны, отсюда следует, что путем должной фиксации величины рН можно избежать выделения белков. Последнее удалось подтвердить экспериментально замораживанием мяса с фиксированным значением рН 6,3, после оттаивания которого потеря сока, характеризующая степень денатурирования белков, оказалась очень малой. 10)Количество вымерзающей воды. Вымерзание воды из растительных и животных клеток начинается при температуре их замерзания. По мере отбора тепла вымерзает все больше воды, а концентрация растворенных в клеточной влаге веществ увеличивается. После достижения определенной точки снижать температуру далее бесполезно, поскольку вода из клеток больше не вымерзает. Некоторое ее количество даже при очень низких температурах остается в жидкой фазе. С помощью современного дифференциального термоанализа Дакуорт (1971) установил количество невымерзающей из продуктов воды при температуре —180° С (табл. ). |


Copyright © 2012 г.
При использовании материалов - ссылка на сайт обязательна.